Spéciation
La spéciation est, en biologie, le processus évolutif par lequel de nouvelles espèces vivantes se forment à partir d'ancêtres communs.
Les espèces s'individualisent à partir de populations appartenant à une espèce d'origine (sauf exception pour certains végétaux). Une espèce, au sens du concept biologique d'espèce, est définie comme ayant une communauté d'ascendance au sein de laquelle tout individu est interfécond avec les autres et donnera une descendance fertile[1]. Il y a donc eu spéciation lorsque deux groupes partageant les mêmes ancêtres ne sont plus interféconds, et prennent ainsi le statut d'espèces biologiques vraies. La "zone grise" dans laquelle deux populations commencent à diverger et au-delà de laquelle on considère que ces deux groupes sont désormais deux espèces distinctes s'étend de 0,5 % jusqu'à 2 % de divergences moléculaires[2].
La spéciation résulte de la dérive génétique et de la sélection naturelle, qui sont les deux moteurs de l'évolution. Une espèce n'apparaît pas instantanément par une mutation conduisant à l'apparition d'un individu d'un type nouveau. Au sein d'une population, les individus varient aléatoirement au cours du temps, c'est le principe de divergence[1]. Charles Darwin fut le premier à décrire le rôle de la sélection naturelle comme facteur de spéciation, dans son livre L'Origine des espèces (1859)[3].
Ce terme a été introduit en 1906 par le biologiste Orator F. Cook pour décrire la séparation d'une lignée ancestrale en clades distincts, ou cladogénèse, par opposition à l'évolution phylétique au sein d'une même lignée, ou anagénèse[4],[5],[6].
Le concept de spéciation a été essentiellement développé par Ernst Mayr.
On distingue quatre modes de spéciations dans la nature, suivant le degré d'isolement d'une sous-population par rapport aux autres : allopatrique (formation d'une barrière physique), péripatrique (par migration), parapatrique (par colonisation d'une niche écologique voisine) et sympatrique (par colonisation d'une niche écologique ou comportementale dans la même aire de répartition). On peut également créer des spéciations artificielles, par élevage, ou par des expériences de laboratoire.

Comparaison des différents modes de spéciation.
Sommaire
1 Mécanisme de spéciation
1.1 Espèce et dérive génétique
1.2 Spéciation et dérive génétique
1.3 Spéciation par opportunité
1.4 Vitesse d'évolution
2 Types de spéciation
2.1 Spéciation allopatrique
2.1.1 Spéciation vicariante
2.1.2 Spéciation péripatrique ou spéciation par effet fondateur
2.1.3 Spéciation parapatrique
2.2 Spéciation sympatrique
2.3 Spéciation par distance
3 Vitesse de spéciation
4 Notes et références
5 Voir aussi
5.1 Vidéo
5.2 Articles connexes
Mécanisme de spéciation |
Espèce et dérive génétique |

Les variations individuelles des génotypes induisent des variations dans les phénotypes, et de là dans les performances individuelles, sources de sélection.
Au niveau génétique, une espèce correspond à un pool génétique, où les différents gènes peuvent exister sous forme de variantes (allèles) ; mais ces différences génétiques n'empêchent pas l'interfécondité entre individus. La reproduction sexuée réalise un brassage génétique qui assure à long terme et en moyenne l'homogénéisation du pool génétique.
La dérive génétique créé constamment de nouveaux allèles, et ainsi entretient une variabilité dans la population. Ces différents allèles peuvent réaliser au niveau individuel des phénotypes légèrement différents, ou au contraire être relativement neutres dans leur expression. Les variations individuelles des phénotypes peuvent entraîner des différences dans la sélection naturelle et le succès reproductif des individus.
Dans ce cas, les allèles statistiquement associés à un moindre succès reproductif (pour une raison ou une autre) seront sélectionnés négativement, et ceux au contraire qui confèrent statistiquement un avantage reproductif verront leur fréquence augmenter dans le pool génétique. L'effet de la sélection naturelle sera donc de stabiliser le pool génétique sur un contenu qui, dans les générations passées, a été statistiquement optimal pour les individus recevant le phénotype associé, puisqu'assurant le meilleur succès reproductif possible. La sélection naturelle tend donc à s'opposer à la dérive génétique, et à réduire la variabilité de la population.
Aussi longtemps que l'écosystème dans lequel se place une espèce est stable, son phénotype sera donc stable, puisqu'il ne peut pas s'écarter significativement de son optimum. Pour autant, son pool génétique dérivera en permanence, sous l'effet de variations neutres.
Si donc une espèce peut être définie à un instant donné par son pool génétique, le phénotype associé, et/ou l'interfécondité effective de ses membres, dans le temps les espèces évoluent continuellement et continument. Même à phénotype constant, la dérive génétique peut être telle que la relation d'interfécondité, évidemment respectée d'une génération à l'autre, n'est plus assurée par transitivité après un trop grand nombre de générations. Et si l'interfécondité n'est plus réalisée entre deux sous-groupes, les pools génétiques finiront par diverger, ainsi probablement que le phénotype.
Spéciation et dérive génétique |
La reproduction sexuée réalise un brassage génétique qui assure en moyenne l'homogénéisation du pool génétique. Une spéciation ne peut donc intervenir que si en amont le brassage génétique cesse d'opérer. Par la suite, la disparition d'interfécondité entre les sous-populations signe l'existence d'espèces à part entière.
- Le cas le plus simple à concevoir est celui de l'isolement écologique par coupure des aires de répartition, mettant fin mécaniquement au brassage génétique. Dans ce cas, les populations isolées évoluent de manière à optimiser le phénotype sur chaque sous-aire de répartition, et la dérive génétique assurera à terme l'isolement gamétique de ces populations indépendantes.
- Un cas plus complexe est celui d'une colonisation de différentes niches écologiques par une même espèce dans une même aire de répartition, comme l'explosion radiative des cichlidae dans les grands lacs africains. Dans ce cas, la colonisation et la spécialisation sur une nouvelle niche écologique (qui correspond à une dérive du phénotype comportemental) doit nécessairement être associée à une sélection sexuelle provoquant un emballement fisherien sur cette sous-population, conduisant à son isolement reproductif.
Dans tous les cas, l'isolement éthologique peut être assuré ou renforcé par non perception des signaux de parade visuels, olfactifs (phéromones, cas des papillons de nuit Yponomeuta) ou auditifs (chants, exemple de Sturnella magna et Sturnella neglecta)[7]. La sélection sexuelle peut être le seul facteur comportemental prévenant l'hybridation de deux sous-populations parfaitement interféconde par ailleurs[Information douteuse] [?][8]. Par la suite, l'interfécondité peut être rendue définitive par l'isolement mécanique et l'isolement gamétique, quand la copulation ne peut avoir lieu ou s'il y a copulation qu'elle ne donne pas de fécondation[9].
Spéciation par opportunité |
La très grande variabilité du vivant dans les plans d'organisation et la complexité des organismes peut donner l'impression que l'évolution procède par additions et complexité croissante, comme le progrès technologique. Mais ce n'est que très rarement le cas. Le répertoire génétique humain ne diffère pas tellement de celui des souris ou des grenouilles, l'évolution vers telle ou telle forme n'a pas consisté à inventer de nouveaux tissus mais à déplacer ou déformer des constructions préexistantes[10]. L'évolution procède par adaptation et diversification : s'il est possible d'occuper une niche écologique à moindre pression sélective par une petite variation de l'existant, cette niche finira toujours par être occupée d'une manière ou d'une autre[10]. Une fonctionnalité nouvelle résulte toujours au départ d'une possibilité accidentellement offerte par une variation quantitative, et qui peut dans un deuxième temps être exprimée et développée qualitativement. L'impression de complexité croissante vient de ce que les systèmes simples sont déjà occupés et saturés, tandis qu'il n'y a pas de limite dans le sens d'une complexité croissante[10].
Un scénario évolutif ne peut pas se limiter à indiquer que tel nouveau phénotype présente un avantage parce qu'il permet telle ou telle possibilité. Il faut préciser quel était au départ le trait primitif d'où ce nouveau trait a évolué, en quoi ce trait primitif a pu être (par lui-même ou dans une partie du domaine couvert par sa variabilité) une pré-adaptation à une faculté nouvelle, susceptible de présenter un avantage sélectif dans un nouveau contexte, et ceci malgré son coût biologique qui empêchait de dériver en ce sens dans le contexte précédent. Ceci étant acquis, la pression de sélection fera alors effectivement évoluer ce trait vers une optimisation croissante de sa faculté, réalisant l'avantage à travers un nouveau phénotype[10].
Vitesse d'évolution |
L'évolution associée à un avantage reproductif peut être très rapide à l'échelle des temps géologiques. Ainsi, si les conditions écologiques font qu'une petite taille est légèrement avantagée par rapport aux grandes, si bien qu'à chaque génération la taille moyenne de la population diminue de un pour mille (ce qui représenterait deux millimètres à l'échelle humaine), il suffit de 700 générations pour que la taille moyenne soit divisée par deux (1.001700≈2.013{displaystyle 1.001^{700}approx 2.013}
Types de spéciation |
On distingue plusieurs types de spéciation, basés sur la répartition géographique des populations en divergence au cours du processus.
Spéciation allopatrique |
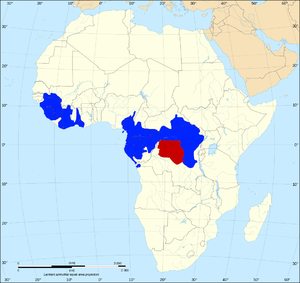
L'ombre rouge indique l'habitat des bonobos. L'ombre bleue indique l'habitat des chimpanzés. C'est un exemple de spéciation allopatrique parce qu'ils sont séparés par une barrière naturelle (la rivière Congo) et ne partagent aucune partie de leur habitat.
Selon ce mode de spéciation, des populations initialement interfécondes évoluent en espèces distinctes car elles sont isolées géographiquement. C'est le mode de spéciation de loin le plus fréquent chez les animaux. Il se décline en deux modalités :
Spéciation vicariante |
Une barrière géographique (rivière, montagne, vallée, océan, glacier…) coupe l'aire de répartition d'une espèce en plusieurs zones. Dans chacune des zones, chaque population évolue indépendamment des autres, pouvant donner naissance à une nouvelle espèce. C'est le cas des éléphants nains comme Elephas falconeri en Sicile.
Spéciation péripatrique ou spéciation par effet fondateur |
Un petit nombre d'individus fondent une nouvelle population en marge de l'aire de répartition de l'espèce d'origine, par exemple à la suite de la colonisation d'une île près de la côte. Cette nouvelle population de petite taille peut évoluer rapidement en une nouvelle espèce.
Ces populations peuvent aussi revenir en sympatrie (contact secondaire), et peuvent même donner une descendance si les évolutions n'ont pas été trop importantes. Dans ce cas, il peut tout de même y avoir une spéciation s'il y a une faiblesse hybride, c'est-à-dire que la survie/reproduction de l'hybride est plus faible (même légèrement) que celle des deux populations. Dans ce cas les allèles conférant la capacité de reproduction avec les individus de l'autre population seront donc désavantagés et disparaîtront. On parle alors de mécanismes de renforcement qui apparaissent dès qu'il y a une amorce de reproduction (faiblesse hybride).
Les espèces Panthera leo et Panthera tigris illustrent parfaitement ce concept. Actuellement, ce sont deux espèces différentes, elles ne peuvent se reproduire l'une avec l'autre qu'en captivité.
Lors du contact secondaire, les lions et les tigres pouvaient donc encore se reproduire, mais les tigrons et les ligres (hybride) présentent une faiblesse (système immunitaire défaillant pour le tigron…). Les lions se reproduisant avec des tigres ont donc eu moins de descendants et ceux-ci ont progressivement disparu. Actuellement aucun lion ne se reproduit avec un tigre en conditions naturelles. Il s'agit bien d'une spéciation.
L'apparition du moustique du métro londonien, la sous-espèce Culex pipiens molestus, est également un exemple de spéciation allopatrique en cours. Au cours des travaux de réalisation des voies du métro londonien, quelques moustiques se sont introduits dans les tunnels souterrains où ils se nourrissent plutôt du sang de mammifères (rats, souris, voyageurs) alors que la population vivant à l'air libre se nourrit préférentiellement de sang d'oiseaux. Durant la Seconde Guerre mondiale, les Londoniens qui s'y réfugiaient lors des bombardements se plaignent des attaques de ces moustiques voraces. En surface, les deux formes de Culex pipiens coexistent et peuvent se reproduire encore entre elles[11].
Dans l'espèce Canis familiaris, on comprendra aisément que la fécondation réussie de femelles chihuahua par des mâles Saint-Bernard pose une double difficulté exclusivement physique :
- la possibilité pratique de l'accouplement,
- celui d'une arrivée de la gestation à bon terme,
même si les gamètes sont compatibles in vitro : la raciation commencerait dans ce cas précis - sans l'existence d'intermédiaires - son cheminement vers une possible spéciation.
A contrario, certaines races de Canis lupus sont interfécondes avec Canis familiaris et les croisements donnent une descendance féconde. Les notions de race et d'espèce contiennent donc parfois une réduction simplificatrice de la complexité du vivant. (voir Classification). Il faut noter qu'actuellement cet exemple ne semble plus pertinent, les chiens (Canis familiaris) étant considérés comme une sous-espèce de Canis lupus sous le nom de Canis lupus familiaris.
Spéciation parapatrique |
Dans un cas de spéciation parapatrique, les populations en divergence ne sont pas totalement isolées géographiquement mais possèdent une zone de contact étroite. Les migrations entre populations sont cependant limitées puisque ces dernières se perpétuent dans des conditions environnementales différentes (gradient climatique par exemple). La sélection naturelle a donc un rôle important dans ce mode de spéciation.
La spéciation parapatrique concerne par exemple les corneilles (Corvus sp.) du paléarctique occidental : on y trouve la Corneille noire (Corvus corone) à l'ouest et la Corneille mantelée (Corvus cornix) à l'est, mais les deux espèces s'hybrident régulièrement dans une zone de contact formant une étroite bande reliant le Danemark, l'est de l'Autriche et le sud des alpes françaises[9].
Spéciation sympatrique |
Des populations non isolées géographiquement peuvent évoluer en espèces distinctes. Ici, la sélection naturelle joue un rôle crucial dans la divergence des populations. Ce phénomène reste controversé, mais semble prouvé chez plusieurs poissons (notamment les cichlidés), ou chez certains insectes phytophages. La différenciation des espèces pourrait aussi résulter du conflit sexuel.
Variante : chez certains végétaux supérieurs (angiospermes), deux espèces normalement non-interfécondes peuvent engendrer des hybrides féconds, sous certaines conditions liées au doublement du nombre de chromosomes présent dans l'hybride formé (diploïdie). Il peut alors lui-même se perpétuer tout en étant incapable de se reproduire avec des individus appartenant aux espèces de ses parents. C'est donc le premier représentant d'une nouvelle espèce, sans faire intervenir un isolement géographique. On parle donc là aussi de spéciation sympatrique, qui semble très fréquente chez les espèces herbacées.
Un exemple plus rare de spéciation sympatrique est la divergence entre « résident » et « transient » chez l'Orque dans le pacifique[12]. Les deux formes habitent le même océan mais ne chassent pas les mêmes proies, n'ont pas les mêmes émissions vocales et ne se reproduisent pas entre eux. Des divergences entre les espèces peuvent aussi provenir des contraintes de micro-habitats contrastés. Ainsi, le putois présente un phénotype rare dit dark qui est directement dépendant des ruisseaux boisés et établit une convergence avec la physionomie du très rare vison d'Europe[13].
Spéciation par distance |
Dans ce cas deux espèces non interfécondes ont des aires de répartition reliées par une aire occupée par une succession de populations interfécondes présentant une variation graduelle des caractères. La spéciation par distance ou par chevauchement circulaire (ring species ou variation clinale) a été étudiée chez le Pouillot verdâtre[14]. Elle se rencontre aussi chez les goélands Larus argentatus et Larus fuscus[15].
L'hypothèse d'une spéciation à distance des néandertaliens est étudiée car il existe un gradient morphologique du Moyen-Orient vers l'Europe occidentale au sein de la population néandertalienne[14],[16].
Vitesse de spéciation |
Gradualisme phylogénétique : l’histoire de l’évolution montre que les spéciations nécessitent généralement plusieurs centaines de milliers d’années, la spéciation est lente, uniforme et graduelle. On parle d'anagénèse dans la transformation d'une espèce entière en une nouvelle espèce et de cladogénèse en cas de séparation nette entre espèce ancestrale et espèce fille[15].
À contrario, certaines nouvelles espèces de souris comme celles de Madère n'ont nécessité qu'entre 500 et 1 000 ans pour apparaître (en fonction de l’origine viking ou portugaise qu’on leur attribue) ce qui représente environ 2 000 à 4 000 générations soit une vitesse extrêmement rapide.
En opposition avec cette vision (« gradualisme phylétique »), la théorie des équilibres ponctués (Stephen Jay Gould, Niles Eldredge) postule que cette histoire comprendrait de longues périodes d'équilibre, ponctuées de brèves périodes de changements importants comme la spéciation ou les extinctions[15],[17].
Notes et références |
« Spéciation », sur jussieu.fr (consulté le 28 juillet 2012)
(en) Camille Roux, Christelle Fraïsse, Jonathan Romiguier, Yoann Anciaux, Nicolas Galtier, Nicolas Bierne, « Shedding Light on the Grey Zone of Speciation along a Continuum of Genomic Divergence », PLOS Biology, vol. 14, no 12, 27 décembre 2016(DOI 10.1101/059790)
Sara Via, « Natural selection in action during speciation », National Academy of Sciences, Washington, D.C., vol. 106, no Suppl 1, 16 juin 2009, p. 9939–9946 (ISSN 0027-8424, PMID 19528641, PMCID 2702801, DOI 10.1073/pnas.0901397106, lire en ligne [PDF])
Berlocher 1998, p. 3
Orator F. Cook, « Factors of species-formation », American Association for the Advancement of Science, Washington, D.C., vol. 23, no 587, 30 mars 1906, p. 506–507 (ISSN 0036-8075, PMID 17789700, DOI 10.1126/science.23.587.506)
Orator F. Cook, « Evolution Without Isolation », University of Chicago Press on behalf of the American Society of Naturalists, Chicago, IL, vol. 42, no 503, novembre 1908, p. 727–731 (ISSN 0003-0147, DOI 10.1086/279001)
« Phéromones et spéciation », sur versailles.inra.fr (consulté le 28 juillet 2012)
Richard Dawkins, The Cichlid's Tale, in The ancestor's tale (Il était une fois nos ancêtres).
« Spéciation », sur ecosociosystemes.fr (consulté le 28 juillet 2012)
Terrence W. Deacon, The symbolic species: the co-evolution of language and the brain. W. W. Norton & Company, 1997. ISBN 0-393-03838-6.
(en) Katharine Byrne & Richard A Nichols, « Culex pipiens in London Underground tunnels: differentiation between surface and subterranean populations », Heredity, no 82, 1999, p. 7–15 (DOI 10.1038/sj.hdy.6884120)
Burden et al.: Resident And Transient-Type Killer Wales SC/56/SM15.
Thierry Lodé "Genetic divergence without spatial isolation in polecat Mustela
putorius populations". J Evol Biol 14:228-236, 2001
« Néenderthal une spéciation à distance », sur hominides.com (consulté le 28 juillet 2012)
« Spéciation », sur univ-lille1 (consulté le 28 juillet 2012)
Dossier pour la science 76, juillet 2012, Anna Degioanni et Sylvana Condemi, Neandertal, Sapiens et Denisova racontés par leurs gènes
Voir article HSP90
Voir aussi |
Vidéo |
vidéo en français (9 min) de Léo Grasset sur la difficulté de définir une espèce.
Articles connexes |
- Évolution
- Bouquet d'espèces
- Écologie du paysage
- Fragmentation écopaysagère
- Insularisation écologique
- Variation clinale
- Conspécificité
- Spéciation par hybridation
- Nouvelles espèces biologiques
 Portail de l’évolution
Portail de l’évolution


